《《DNA的復(fù)制和修復(fù)》PPT課件》由會(huì)員分享���,可在線閱讀��,更多相關(guān)《《DNA的復(fù)制和修復(fù)》PPT課件(40頁珍藏版)》請(qǐng)?jiān)谘b配圖網(wǎng)上搜索����。
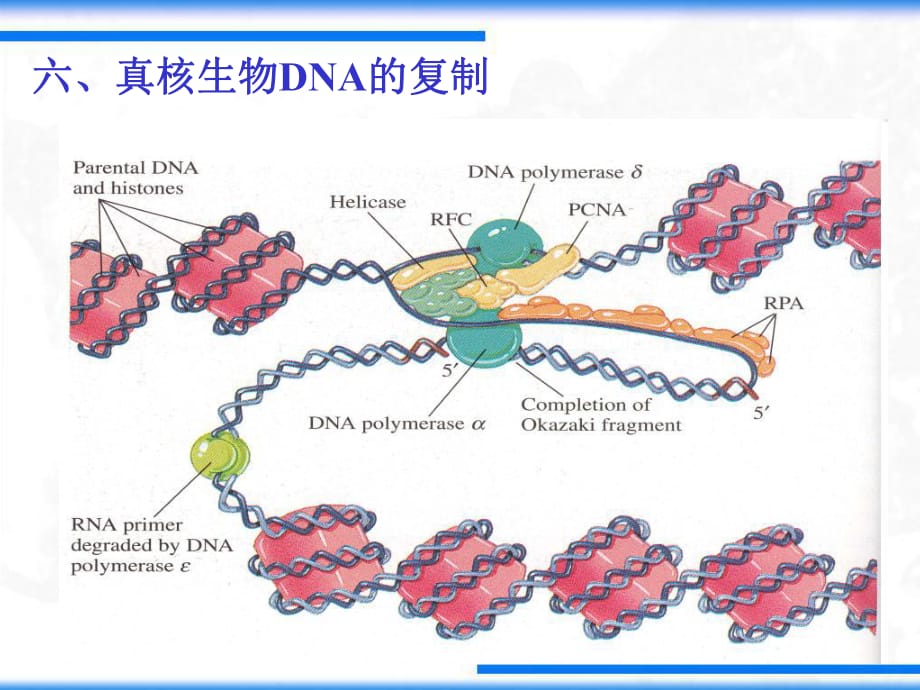
1����、六����、真核生物DNA的復(fù)制 (一)參與復(fù)制的酶及蛋白質(zhì)因子1. DNA聚合酶:pol ����、 、 (I) (I V) (M) (I I I) (I I)位置細(xì)胞核細(xì)胞核線粒體細(xì)胞核細(xì)胞核亞基數(shù)目4 1 2 2 13 5 外切酶- - + + +引物合成酶+ - - - -持續(xù)合成能力中等低高有PCNA時(shí)高高準(zhǔn)確性高低高高高 功能引物合成修復(fù)線粒體DNA合成核DNA合成修復(fù) 2. 復(fù)制體的組成(與原核生物相比較)真 核 生 物復(fù) 制 酶 pol/ 進(jìn) 行 性 因 子 增 殖 細(xì) 胞 核 抗 原 ( PCNA)夾 子 裝 置 器 RF-C(replication factor C)引 物 合 成 酶 p
2����、ol去 除 引 物 RNaseH1 和 MF-1( 53外 切 酶 )滯 后 鏈 修 復(fù) pol 和 DNA連 接 酶消 除 拓 撲 張 力 拓 撲 異 構(gòu) 酶解 螺 旋 酶 T抗 原單 鏈 結(jié) 合 RP-A ( replication protein A) SSBDnaB旋 轉(zhuǎn) 酶pol和 DNA連 接 酶polDnaG復(fù) 合 物夾 子pol 原 核 生 物 A schematic view of the major components at the eukaryotic replication fork.MF-1(單鏈結(jié)合蛋白)(復(fù)制酶)(夾子)(夾子裝置器)(拓?fù)洚悩?gòu)酶)(解旋酶)(引
3、物合成酶)(DNA連接酶) Structure of the PCNA homotrimer. Note that the trimeric PCNA ring of eukaryotes is remarkably similar to its prokaryotic counterpart, the dimeric sliding clamp. (二)真核生物DNA復(fù)制過程1.真核與原核生物在復(fù)制過程上的相同之處: -半 不 連 續(xù) 復(fù) 制 ����; -具 有 相 似 的 起 始 、 延 伸 等 過 程 ��。 原 核 生 物 真 核 生 物復(fù) 制 子 1個(gè) 多 個(gè)起 始 點(diǎn) 1個(gè) ( 可 連 續(xù)
4��、啟 動(dòng) 復(fù) 制 ) 多 個(gè)延 伸 速 度 105bp/min 10003000bp/min岡 崎 片 段 10002000個(gè) 核 苷 酸 100200個(gè) 核 苷 酸引 發(fā) 策 略 引 物 合 成 酶 催 化 合 成 引 物 pol催 化 合 成 引 物 和寡 聚 脫 氧 核 苷 酸延 伸 策 略 由 pol異 源 二 聚 體 催 化 前 導(dǎo)鏈 和 滯 后 鏈 的 合 成 兩 個(gè) pol分 別 催 化 前 導(dǎo)鏈 和 滯 后 鏈 的 合 成2. 真核與原核生物DNA復(fù)制一般特點(diǎn)的比較 (三)端粒及端粒的合成1. 端粒(telomere) 真 核 生 物 線 性 染 色 體 末 端 的 特 殊 結(jié)
5�、構(gòu) , 由 許多 成 串 短 的 重 復(fù) 順 序 組 成 ����, 具 有 穩(wěn) 定 染 色 體 末端 結(jié) 構(gòu) 的 功 能 �。四 膜 蟲 TTGGGG(僅 列 一 條 鏈 的 序 列 )人 TTAGGG 2. 端粒酶(telomerase) 含 有 RNA鏈 的 逆 轉(zhuǎn) 錄 酶 ����, 它 以 所 含 RNA為 模板 來 合 成 DNA端 粒 結(jié) 構(gòu) 。3. 端粒的合成 端 粒 酶 結(jié) 合 到 端 粒 的 3末 端 上 �����, 酶 分 子 中RNA鏈 5 末 端 識(shí) 別 DNA 3末 端 并 互 補(bǔ) 配 對(duì) ����。 以RNA鏈 為 模 板 使 DNA鏈 延 伸 , 合 成 一 個(gè) 重 復(fù) 單位 后 再 向 前 移
6�����、 動(dòng) 一 個(gè) 單 位 �。 3單 鏈 可 回 折 作 為引 物 �, 合 成 其 互 補(bǔ) 鏈 。 (a)Telomeres on human chromosomes consist of the hexanucleotide sequence TTAGGG repeated between 1000 and 1700 times. (b) Like other telomerases, human telomerase is a ribonucleoprotein. Problem posed in the replication of linear DNA : the end of one da
7�����、ughter strand will be shortened after each round of replication. Synthesis of telomeric DNA by telomerase extends the 3-end. 七���、生物體DNA復(fù)制的基本特點(diǎn) 復(fù) 制 方 式 為 半 保 留 復(fù) 制 ����。 復(fù) 制 起 始 于 特 定 位 置 ; 原 核 生 物 ����、 病 毒 是 單 復(fù) 制子 , 具 一 個(gè) 復(fù) 制 起 點(diǎn) ����; 真 核 生 物 是 多 復(fù) 制 子 , 具 多個(gè) 復(fù) 制 起 點(diǎn) �����。 復(fù) 制 有 單 向 和 雙 向 復(fù) 制 ( 后 者 更 為 常 見 ) ��。 復(fù) 制
8���、 時(shí) ���, DNA鏈 由 53延 伸 , 需 RNA引 物 ����, 復(fù) 制 過 程 為 半 不 連 續(xù) 復(fù) 制 �����。 第二節(jié) DNA的損傷及修復(fù) 導(dǎo)致DNA損傷的因素生物因素:物理因素:化學(xué)因素:復(fù) 制 錯(cuò) 誤 等 ��。紫外線電離輻射如:HNO2 烷化劑可 使 同 一 條 鏈 上 相 鄰 嘧 啶 形 成嘧 啶 二 聚 體(TT, CT, CC) 打 斷 DNA雙 鏈 或 單 鏈 �, 破 壞 堿 基使 堿 基 脫 氨 基 �����, AI, CU�, 造 成堿 基 錯(cuò) 配可 導(dǎo) 致 堿 基 錯(cuò) 配 、 DNA交 聯(lián) ��、 堿 基脫 嘌 呤 等 修復(fù)系統(tǒng) 錯(cuò) 配 修 復(fù) 直 接 修 復(fù) 切 除 修 復(fù) 重 組 修 復(fù)
9�、傾 錯(cuò) 修 復(fù) (一)錯(cuò)配修復(fù)(mismatch repair)定 義 : 復(fù) 制 后 的 DNA在 短 時(shí) 間 內(nèi) GATC序 列 是 半 甲 基化 的 ���, 一 旦 發(fā) 現(xiàn) 錯(cuò) 配 堿 基 �����, 包 括 錯(cuò) 配 堿 基 在 內(nèi) 的 未甲 基 化 的 新 鏈 可 被 切 除 ���, 并 以 甲 基 化 的 鏈 為 模 板 進(jìn)行 修 復(fù) 合 成 �����。 (二)直接修復(fù)某 些 損 傷 的 核 苷 酸 和 錯(cuò) 配 的 堿 基 可 以 被 某 些蛋 白 質(zhì) 識(shí) 別 和 修 復(fù) ��, 這 些 蛋 白 質(zhì) 為 了 能 夠 找出 特 別 損 傷 部 位 可 以 連 續(xù) 監(jiān) 測(cè) DNA���。 這 些 蛋白 質(zhì) 不 切 斷 D
10、NA或 切 除 堿 基 而 是 直 接 實(shí) 施 修復(fù) ����, 這 樣 的 損 傷 修 復(fù) 機(jī) 制 稱 為 直 接 修 復(fù) 。 可 見 光 激 活 光 復(fù) 活 酶 ���, 其 可 以 分 解由 于 紫 外 線 照 射 形 成 的 嘧 啶 二 聚 體 ��, 恢復(fù) DNA的 正 常 結(jié) 構(gòu) ����。光復(fù)活修復(fù)(photoreactivation repair) 此 修 復(fù) 方 式 從 低 等 生 物 到 鳥 類 皆 有 �����,在 植 物 中 比 較 重 要 。 DNA is distorted at the site of T-T dimer. 通 過 光 復(fù) 活 酶 修 復(fù) 胸 腺 嘧 啶 二 聚 體 的 過 程 (
11����、三)切除修復(fù)(excision repair)v定義:v 包括:堿 基 切 除 修 復(fù) ( base-excision repair)核 苷 酸 切 除 修 復(fù) ( nucleotide-excision repair) 在 一 系 列 酶 的 作 用 下 , 將 DNA分 子 中 受 損 傷部 分 切 除 掉 ��, 并 以 完 整 鏈 的 一 條 鏈 為 模 板 ���, 合 成切 去 的 部 分 ��, 使 DNA恢 復(fù) 正 常 結(jié) 構(gòu) 的 過 程 ���。 (四)重組修復(fù)v定義: 含 有 嘧 啶 二 聚 體 或 其 他 結(jié) 構(gòu) 損 傷 的 DNA進(jìn) 行 復(fù) 制時(shí) , 子 代 DNA鏈 在 損 傷 部 位
12����、出 現(xiàn) 缺 口 。 通 過 遺 傳 重 組 ���,從 完 整 的 母 鏈 上 將 相 應(yīng) 的 片 段 移 至 子 鏈 的 缺 口 處 ��, 然后 用 再 合 成 的 多 核 苷 酸 鏈 補(bǔ) 上 母 鏈 的 空 缺 。 (五)應(yīng)急反應(yīng)(SOS)和易錯(cuò)修復(fù) 許 多 可 造 成 DNA損 傷 或 抑 制 復(fù) 制 的 處 理 均 能 引起 一 系 列 復(fù) 雜 的 誘 導(dǎo) 反 應(yīng) ����, 稱 為 SOS反 應(yīng) ����。 1. 應(yīng)急反應(yīng)包括:DNA損 傷 修 復(fù)誘 變 效 應(yīng)細(xì) 胞 分 裂 的 抑 制溶 原 性 細(xì) 菌 釋 放 噬 菌 體( 細(xì) 胞 癌 變 也 與 SOS反 應(yīng) 有 關(guān) ) 傾 錯(cuò) 的 DNA聚 合 酶
13���、IV����、 V避免差錯(cuò)的修復(fù)(error free repair)傾向差錯(cuò)的修復(fù)(error prone repair)2. SOS反應(yīng)誘導(dǎo)的修復(fù)系統(tǒng)錯(cuò) 配 修 復(fù) �、 直 接 修 復(fù) 、切 除 修 復(fù) ���、 重 組 修 復(fù)3. SOS反應(yīng)機(jī)制 由Rec A蛋白和LexA阻遏物相互作用引起的��。 4. SOS反應(yīng)的意義 SOS反 應(yīng) 包 括 兩 方 面 : DNA修 復(fù) 和 導(dǎo) 致 變異 ����。 修 復(fù) 可 使 DNA恢 復(fù) 正 常 結(jié) 構(gòu) �����, 而 變 異 可 增加 物 種 生 存 的 機(jī) 會(huì) , 在 進(jìn) 化 中 是 十 分 有 意 義 的 �����。 第三節(jié) RNA指導(dǎo)下的DNA的合成 逆轉(zhuǎn)錄作用(revers
14�、e transcription) 是 以 RNA為 模 板 , 即 按 照 RNA中 的核 苷 酸 順 序 合 成 DNA的 過 程 �。 1964 年 , Temin 提 出 前 病 毒 學(xué) 說 ��, 即 在 致 癌RNA病 毒 的 感 染 過 程 中 存 在 一 個(gè) DNA中 間 體( 前 病 毒 ) ����。 1970年 , Temin和 Baltimore分 別 從 致 癌 RNA病毒 中 發(fā) 現(xiàn) 逆 轉(zhuǎn) 錄 酶 �。(一)逆轉(zhuǎn)錄酶(reverse transcriptase)的發(fā)現(xiàn)致 癌 RNA病 毒 是 一 大 群 能 引 起 鳥 類 、 哺 乳 類 等 動(dòng) 物 白 血 病 和肉 瘤 以 及
15�����、其 它 腫 瘤 的 病 毒 ����。 這 類 病 毒 侵 染 細(xì) 胞 后 并 不 引 起 細(xì)胞 死 亡 , 卻 可 以 使 細(xì) 胞 發(fā) 生 惡 性 轉(zhuǎn) 化 �����。 1975年 ��, 獲 諾貝 爾 生 理 醫(yī) 學(xué)獎(jiǎng) ���。 (二)逆轉(zhuǎn)錄酶的性質(zhì)活性: RNA指導(dǎo)的DNA聚合酶活力 DNA指導(dǎo)的DNA聚合酶活力 核糖核酸酶H的活力以 4種 dNTP為 底 物需 要 有 模 板 和 引 物聚 合 方 向 : 53 需 Mg2+( Mn2+) 數(shù)目:70個(gè)/ 病毒粒子 結(jié)構(gòu):含����、兩個(gè)亞基 逆轉(zhuǎn)錄病毒的基因組通常有兩條相同的(+)RNA鏈所組成�����,在靠近5端的區(qū)域以氫鍵結(jié)合在一起���。 5端具有帽子結(jié)構(gòu)��,3端具有多聚腺苷酸����。
16��、靠近5有1分子宿主tRNA���,作為逆轉(zhuǎn)錄引物���。典型逆轉(zhuǎn)錄病毒基因組長(zhǎng)約7000-100000核苷酸����,攜帶3個(gè)基因(gag�,編碼核心蛋白、pol��,編碼蛋白酶�����、整合酶����、逆轉(zhuǎn)錄酶、env�,編碼膜蛋白)?��;蚪MRNA的兩端具有同樣的序列�,成為正向重復(fù)�。 General structure of an integrated retrovirus genome. The long terminal repeats (LTRs) have sequences needed for the regulation and initiation of transcription. The sequence deno
17�����、ted is required for packaging retroviral RNAs into mature virus particles. (三)逆 轉(zhuǎn) 錄 病 毒 生 活 史 :逆 轉(zhuǎn) 錄 病 毒在 逆 轉(zhuǎn) 錄 酶 的 催 化 下 �, 以 病 毒 的 RNA為 模 板 合 成�, 形 成 RNA-DNA雜 化 體 ��, 逆 轉(zhuǎn) 錄 酶將 雜 化 體 中 的 RNA降 解 �, 同 時(shí) 以 剩 下 的 DNA鏈 為 模 板合 成 另 一 條 互 補(bǔ) 的 DNA鏈 ,雙 鏈 DNA 中利 用 宿 主 復(fù) 制 和 轉(zhuǎn) 錄 機(jī) 器 生 產(chǎn) 大 量 的 轉(zhuǎn) 錄 出 的 mRNA翻 譯 成 病 毒
18�、的 包 膜 蛋 白 , 逆 轉(zhuǎn) 錄 酶和 殼 體 蛋 白將 病 毒 RNA��、 逆 轉(zhuǎn) 錄 酶 和 殼 體 蛋 白 組 裝 成 病 毒 的核 殼 體核 殼 體 結(jié) 合 包 膜 蛋 白 形 成逆 轉(zhuǎn) 錄 酶 沒 有 3-5外 切 酶 活 性 或 校 正 活 性 ��, 所 以 它的 錯(cuò) 誤 率 比 任 何 DNA聚 合 酶 都 高 +病毒RNA侵染宿主細(xì)胞RNA-cDNA雜交分子RNA指導(dǎo)的DNA聚合酶-DNA整合入染色體基因組DNA指導(dǎo)的DNA聚合酶- +雙鏈DNA逆轉(zhuǎn)錄過程核糖核酸酶H病毒RNA 自身蛋白質(zhì)轉(zhuǎn)錄翻譯+ cDNA: 幾 乎 所 有 真 核 生 物 mRNA分 子 的 3 末 端 都 有一 段 polyA,當(dāng) 加 入 寡 聚 dT作 為 引 物 時(shí) ��, mRNA就 可 作為 模 板 ��, 在 逆 轉(zhuǎn) 錄 酶 催 化 下 在 體 外 合 成 與 其 互 補(bǔ) 的DNA����, 稱 為 cDNA。 (四)逆轉(zhuǎn)錄酶應(yīng)用與逆轉(zhuǎn)錄研究意義逆轉(zhuǎn)錄酶的應(yīng)用(如:cDNA的獲得�、逆轉(zhuǎn)錄病毒病毒作為信息載體作為基因治療等)研究意義:揭示HIV病毒和嗜肝DNA病毒的致病機(jī)理與防治研究用于分子生物學(xué)研究的材料
 《DNA的復(fù)制和修復(fù)》PPT課件
《DNA的復(fù)制和修復(fù)》PPT課件